Aleochara (Coprochara)
Christian Maus and James S. Ashe (1947-2005)

.png?x=-827289361)

This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
Coprochara is one of the few subgenera in the present subgeneric system of Aleochara which are well-supported monophyletic lineages. Twenty-eight species are currently described (one doubtful species is indicated with a "?" in the species list). Coprochara occurs in all biogeographic regions except the Australian and the Antarctic Region. In the Oriental Region, only few species, which are of mainly Palaearctic distribution, occur in India and China. Because of external similarity, the extraordinary infraspecific variability of the species, and the incorrectly assumed similarity of their male genitalia, Coprochara has always been judged to be one of the most difficult groups within Aleochara.
Characteristics
Autapomorphies of Coprochara are: 1) two longitudinal, parallel or subparallel rows of punctures on the pronotum, whose interspace along the pronotal midline is glabrous ("dorsal rows"); 2) duct of female spermatheca coiled around a virtual longitudinal axis (which is sometimes curved); number of coils from one up to over one hundred (probably secondarily reduced in two species lacking coils);
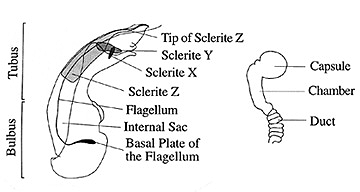
Figure 1. Generalized aedaegus and spermatheca of Coprochara species.
3) median lobe of aedeagus equipped with characteristically arranged sclerites, which can be assigned to two or three different types, respectively (Sclerites X, Y and Z, classification after KLIMASZEWSKI 1984). A further important feature of the subgenus Coprochara is the carinate mesosternum, which is a synapomorphy that it shares with some other subgenera (KLIMASZEWSKI 1984).
The species are small to medium-sized (ca 1.5-10 mm) and usually have a cylindrical, subcylindrical or flattened subparallel body shape. Most species are black or blackish brown, many with a yellow or red terminal spot on each elytron or with indistinctly lightened elytral apex.
Identification Guides
Recent identification keys for Coprochara species are available for North America (KLIMASZEWSKI 1984) and Southern Africa (KLIMASZEWSKI & JANSEN 1994). There are also keys for the Central European (LOHSE 1989, as a supplement to LIKOVSKY 1974) and for the British species (WELCH 1997). The key of LOHSE 1989 (in combination with LIKOVSKY 1974) can be used for material from nearly all of Europe, since they include all European species except two endemics from Madeira and the Azores, respectively. Additional characteristics for species identification are given by MAUS in prep. European keys published before 1986 (e.g. PALM 1972, LIKOVSKY 1974) are no longer up to date for most species and should only be applied for identification of Coprochara species in combination with current supplements.
Bionomics
As far as known, most Coprochara species live mainly in mammal droppings, but occasionally also on carrion and decaying plant material. Members of one specialized group of three species (A. (C.) salsipotens, A. (C.) sulcicollis and A. (C.) squalithorax) are restricted to ocean shores. Hosts and development of ten species are known (MAUS et al. in prep.). Larvae of these known Coprochara species pupate inside the host puparium.
Several species of this subgenus are of considerable importance as biological control agents. Their host ranges include anthomyiid flies which are serious vegetable pests, such as Delia radicum, D. antiqua etc. This applies in particular to A. (C.) bilineata, A. (C.) bipustulata and A. (C.) verna. MAUS et al. (in prep.) provide a compilation of host records.
Discussion of Phylogenetic Relationships
Until now, the only attempt to reconstruct the phylogeny of Coprochara has been made by MAUS 1996 (in an unpublished M.S.thesis). KLIMASZEWSKI & JANSEN (1994) provide a classification that is not based on cladistic principles. The discussion below is adapted from MAUS 1996.
The examination of phylogenetic relationships within Coprochara is very difficult due to the high frequency of parallelism and homoplasy in this group. Most of the apomorphic character states have been developed independently in different lineages during evolution of Coprochara. The cladogram provided here must be considered as provisional; some points must be critically evaluated, and further investigations may result in changes in the cladogram. Since the phylogenetic resolution provided by morphological characteristics is apparently limited for this group, molecular systematic studies are currently under way by one of us (Ch. Maus).
A. (C.) squalithorax, which has previously been assigned to the monotypic subgenus Skenochara (BERNHAUER & SCHEERPELTZ 1926, ASSING 1995), has all apomorphies that characterize Coprochara. The distinctly impressed pronotal dorsal rows and the strongly coiled spermathecal duct of A. (C.) squalithorax suggest that it is a highly derived species in the context of this subgenus (see below). Its exclusion from Coprochara would make the rest of the subgenus paraphyletic, so it is here included in Coprochara (see MAUS in prep.).
In the following discussion the most frequently mentioned character states, which are considered to be apomorphic, are: cylindrical or flattened body shape (in contrast to a rather fusiform, convex body shape); strongly impressed pronotal dorsal rows; elytral pubescence directed almost straight posteriad (in contrast to pubescence directed to the posterior outer angles of the elytra); rasp-like elytral punctures; a large-sized aedeagal sclerite Z; and, spermathecal duct with numerous coils.
A. (C.) densissima is considered to represent the most basal evolutionary lineage within Coprochara due to the more or less fusiform body shape, relatively dense pronotal pubescence and punctation, and other plesiomorphic character states. The other species show less dense pubescence and punctation of the pronotum, features which are interpreted as synapomorphic for the remaining species.
Of the three main lineages, those descending from nodes A and B are made up of mostly rather small, mainly Old-World species, and the lineage descending from node C is made up of rather medium-sized and large species, the majority of which live in America. The first lineage (node A) is characterized by a more or less flattened body, while the members of the second lineage (node B) share the synapomorphy of a rather cylindrical or subcylindrical body shape. The third lineage (node C) is characterized by the presence of a unique third sclerite type (sclerite X) in the male aedeagal median lobe (wich is probably secondarily reduced in some species within this lineage).
========================= A. (C.) densissima
|
| ============ A. (C.) bipustulata
| |
| |=========== A. (C.) lindbergi
| ===|
| | |=========== A. (C.) bisolata
| | |
| | ============ A. (C.) verna
| === A =|
| | | ============ A. (C.) freyi
| | | |
| | | | ========= A. (C.) reinigi
| | ===| |
| | ===| ====== A. (C.) suffusa
| | ===|
| | ====== A. (C.) brundini
| |
| | ============ A. (C.) binotata
| | ===|
| | | ============ A. (C.) pauxilla
| | |
=====| | | ========= A. (C.) hluhluwe
===|== B =| ===|
| | | ========= A. (C.) lineatocollis
| ===|
| | ========= A. (C.) sublaevipennis
| ===|
| ========= A. (C.) granulicauda
|
| =============== A. (C.) bilineata
| |
| | ============ A. (C.) notula
| | |
| | | ====== A. (C.) bimaculata
| | | |
| | | ===| === A. (C.) mutare
=== C =| | | ===|
===| | === A. (C.) solieri
===|
| ====== A. (C.) salsipotens
| |
===| === A. (C.) sulcicollis
===|
=== A. (C.) squalithorax Among the species in the first lineage (node A), four (A. (C.) bipustulata - A. (C.) verna) bear a distinct elytral terminal spot (secondarily reduced in A. (C.) lindbergi), which is a synapomorphy among them (shared in parallel with some species in the third lineage, and indistinct terminal spots are found in other species as well). Among these species, A. (C.) verna is distinct by a large aedeagal sclerite Z. For the remaining complex of three species, no synapomorphies could be found. It is well possible that A. (C.) bipustulata and A. (C.) lindbergi are adelphotaxa due to their extreme similarity and biogeographical considerations that support the assumption that A. (C.) lindbergi is the vicariant species of A. (C.) bipustulata in Madeira, and that their evolutionary lineages have segregated relatively recently. The second complex within this sublineage (A. (C.) freyi - A. (C.) brundini) includes four species which share the synapomorphies of elytral pubescence that is directed almost straight posteriad and rasp-like elytral punctation. Within this complex, A. (C.) suffusa and A. (C.) brundini share the synapomorphy of an increased number of coils of the spermathecal duct, A. (C.) reinigi and the former two species have a large aedeagal sclerite Z as synapomorphy.
The second main lineage (node B), comprising the rather cylindrical species, can be divided into the binotata group (comprising A. (C.) binotata and A. (C.) pauxilla) which is characterized by the rather distinctly impressed pronotal dorsal rows and an increased number of coils of the spermathecal duct, and a complex of four species (A. (C.) hluhluwe - A. (C.) granulicauda) that share the derived condition of a conspicuously slender body shape. Within this complex, the lineatocollis group (comprising A. (C.) hluhluwe and A. (C.) lineatocollis) shows an even more slender body shape, while the sublaevipennis group (comprising A. (C.) sublaevipennis and A. (C.) granulicauda) has secondarily reduced spermathecal coils and aedeagal inner sclerites.
In the third main lineage (node C), all taxa other than the monotypic bilineata group share the synapomorphy of a probably unique pattern of head punctation (with antero-lateral impunctate areas) (secondarily reduced in A. (C.) sulcicollis and A. (C.) aqualothorax. Among these the monophyletic notula group (defined by genitalic features) lacks the apomorphic rather cylindrical body shape shared by the remaining species. The bimaculata group (A. (C.) bimaculata - A. (C.) solieri) is very distinct by synapomorphy of a long aedeagal virga which is coiled in the bulb of the median lobe. In this group, A. (C.) mutare and A. (C.) solieri share the apomorphy of impressed pronotal dorsal rows. The sulcicollis group, which comprises the remaining three species (A. (C.) salsipotens - A. (C.) squalithorax), is characterized by the very strongly impressed pronotal dorsal rows (indistinct in A. (C.) squalithorax) and the seashore habitat. The species of this group lack an aedeagal sclerite X (in A. (C.) sulcicollis, there is a structure which could possibly be interpreted as a rudimentary sclerite X), a condition which is assumed to be a secondary reduction. Within this group A. (C.) sulcicollis and A. (C.) squalithorax are considered to be adelphotaxa because they share a strongly increased number of coils of the spermathecal ductus and emarginate posterior outer angles of the elytra.
The main weakness of this cladogram is inclusion of the sulcicollis group to the lineage defined by presence of three aedeagal inner sclerites. This requires the assumption of a secondary loss of sclerite X within the sulcicollis group. This set of relationships was proposed because of the distinctive punctation pattern of the head (see above), which is the same in A. (C.) salsipotens, and in the notula and bimaculata groups (The unique and highly derived condition of head punctures in A. (C.) sulcicollis and A. (C.) squalithorax is believed to be derived from this condition).
In addition, further characteristics should be evaluated to verify the branches in the tree which are only supported by body shape characteristics.
References
Assing, V. 1995. The Palaearctic species of Emplenota Casey, Polystomota Casey, Triochara Bernhauer and Skenochara Bernhauer & Scheerpeltz, with descriptions of three new species. Beitraege zur Entomologie 45(1): 217-237
Bernhauer, M. & O. Scheerpeltz 1926. Staphylinidae VI. In: Junk, W. & S. Schenkling: Coleopterorum Catalogus. Pars 82. Junk, Berlin. pp. 499-988
Klimaszewski, J. 1984. A revision of the genus Aleochara Gravenhorst in America north of Mexico (Coleoptera: Staphylinidae, Aleocharinae). Memoirs of the Entomological Society of Canada 129: 1-211
Klimaszewski, J. & R. Jansen 1994. Systematics, biology and distribution of Aleochara Gravenhorst from Southern Africa. Part 3: subgenus Coprochara Mulsant and Rey (Coleoptera: Staphylinidae). Annals of the Transvaal Museum 36(10): 147-170
Likovsky, Z. 1974. 237. Gattung: Aleochara Gravenhorst 1802. In: Freude, H., K.W. Harde & G.A. Lohse: Die Kaefer Mitteleuropas. Band 5. Staphylinidae II (Hypocyphtinae und Aleocharinae), Pselaphidae. Goecke & Evers, Krefeld: 381 pp.
Lohse, G.A. 1989. 23. Familie Staphylinidae (II) (Aleocharinae). In: LOHSE, G.A. & W.H. LUCHT: Die Kaefer Mitteleuropas. Band 12 (1. Supplementband). Goeckke & Evers, Krefeld: 346 pp.
Maus, Ch. 1996. Taxonomische und phylogenetisch-systematische Untersuchungen zur Untergattung Coprochara Mulsant & Rey 1874 der Gattung Aleochara Gravenhorst 1802 (Coleoptera, Staphylinidae). M.S. Thesis, Albert-Ludwigs University, Freiburg: 171 pp.
Maus, Ch. in prep.. Taxonomical Contributions to the Subgenus Coprochara Mulsant & Rey, 1874 of the Genus Aleochara Gravenhorst, 1802 (Coleoptera: Staphylinidae).
Maus, Ch., B. Mittmann & K. Peschke. in prep. Survey of host records of parasitoid Aleochara species (Coleoptera: Staphylinidae) attacking cyclorrhaphous fly puparia: Review and new investigations.
Palm, Th. 1972. Skalbaggar. Coleoptera. Kortvingar. Fam. Staphylinidae, Underfam. Aleocharinae (Aleuonota-Tinotus). Svensk Insektfauna 9(7): 460 pp.
Title Illustrations

| Scientific Name | Aleochara bipustulata |
|---|---|
| Reference | Reitter, Edmund. 1908. Die Käfer des Deutschen Reiches. Band 2. Fauna Germanica. K. G. Lutz Verlag, Stuttgart. |
| Source Collection | BioLib Online Library of Biological Books |
About This Page
Development of this page made possible by National Science Foundation PEET grant DEB 95-21755 to James S. Ashe and a DAAD grant from the German Government to Christian Maus.
Photographic images on this page copyright © 1997 James S. Ashe. Drawings of aedeagus and spermatheca of Coprochara copyright © Christian Maus.

Bayer CropScience AG, Institute for Ecotoxicology, Monheim, Germany
James S. Ashe (1947-2005)
University of Kansas, Lawrence, Kansas, USA
Correspondence regarding this page should be directed to Christian Maus at
Page copyright © 1997 and James S. Ashe (1947-2005)
 Page: Tree of Life
Aleochara (Coprochara).
Authored by
Christian Maus and James S. Ashe (1947-2005).
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Aleochara (Coprochara).
Authored by
Christian Maus and James S. Ashe (1947-2005).
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- First online 11 September 1998
Citing this page:
Maus, Christian and James S. Ashe (1947-2005). 1998. Aleochara (Coprochara). Version 11 September 1998 (under construction). http://tolweb.org/Aleochara_%28Coprochara%29/9913/1998.09.11 in The Tree of Life Web Project, http://tolweb.org/