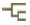Gnathostomata
Jawed Vertebrates
Philippe Janvier


This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxThe main characteristics supporting the nodes of this phylogeny are:
- Node 1: Superior oblique muscle of eye attached anteriorly to eyeball. Braincase including nasal capsules. Jaw muscle external to mandibular arch.
- Teleostomi: Mouth terminal in position, narrow-based braincase, three otoliths in ear.
- Osteichthyes: Endochondral bone, lepidotrichs on fins, jaws lined by dentary, premaxillary and maxillary.
Introduction
The Gnathostomata, or gnathostomes, are the majority of the Middle Devonian (-380 million years ago) to Recent vertebrates. They differ from all other craniates or vertebrates in having a vertically biting device, the jaws, which consist of an endoskeletal mandibular arch and a variety of exoskeletal grasping, crushing, or shearing organs, i.e. the teeth, and jaw bones. Among Recent vertebrates, the gnathostomes include sharks, rays, chimaeras, ray-finned fishes, lobe-finned fishes and land vertebrates.
Extant gnathostomes fall into two major clades, the Chondrichthyes and Osteichthyes. In addition, there are two extinct major gnathostome clades, the Placodermi (Early Silurian-Late Devonian) and the Acanthodii (Latest Ordovician or Earliest Silurian - Early Permian). There may be other fossil gnathostome taxa which fall outside of these four taxa. This could be the case for the Mongolepida, only known from isolated scales from the Early Silurian, and which are provisionally assigned to the chondrichthyans, yet with great reservations.
The Chondrichthyes are characterized by a special type of hard tissue lining the cartilages of the endoskeleton: the prismatic calcified cartilage. Another chondrichthyan characteristic is the pelvic clasper, as special copulatory organ derived from the metapterygium, i.e. the posterior part of the pelvic fin. A pelvic clasper may, however, be present in the fossil Placodermi. Chondrichthyans include two major extant clades, the Elasmobranchii and the Holocephali, and a number of fossil clades (Cladoselachidae, Symmoriida, Xenacanthiformes, Iniopterygia, Eugeneodontida) which may fall outside these two clades.
The Osteichthyes are characterized by endochondral ("spongy") bone in the endoskeleton, dermal fin rays made up by lepidotrichiae (modified, tile-shaped scales), and three pairs of tooth-bearing dermal bones lining the jaws (dentary, premaxillary and maxillary). The Osteichthyes include two major clades, the Actinopterygii and the Sarcopterygii.
The Placodermi are characterized by a dermal armor consisting of a head armor and a thoracic armor. In the thoracic armor, the foremost dermal plates form a complete "ring" around the body and always include at least one median dorsal plate.
The Acanthodii are characterized by dermal spines inserted in front of all fins but the caudal one. They also possess minute, growing scales which have a special onion-like structure, i.e. the crown consists of overlying layers of dentine or mesodentine.
Characteristics
Gnathostomes are characterized by:- A vertically biting device called jaws, and which is primitively made up by two endoskeletal elements, the palatoquadrate and Meckelian cartilage, and a number of dermal elements called teeth, sometimes attached to large dermal bones.
- Pelvic fins. These are the paired fins or limbs situated just in front of the anus.
- Interventrals and basiventrals in the backbone. These are the elements of the backbone which lie under the notochord, and match the basidorsals and interdorsals respectively.
- Gill arches which lie internally to the gills and branchial blood vessels, contrary to the gill arches of all jawless craniates, which are external to the gills and blood vessels.
- A horizontal semicircular canal in the inner ear.
- Paired nasal sacs which are independent from the hypophysial tube. In all extant and fossil jawless craniates, the nasal sacs, which contain the olfactory organs, open into a median duct, the nasohypophysial duct, which takes part to the formation of the pituitary gland and either leads postriorly to the pharynx (e.g. in hagfish and galeaspids) or ends as a blind pouch (e.g. in lampreys and osteostracans). In the gnathostomes, this pouch remains as a thin canal in the palate, the buccohypophysial canal, whereas the nasal sacs open separately to the exterior by external nostrils.

The skull of a gnathostome, or jawed vertebrates (here a shark), are characterized by vertically biting jaws (red) consisting of the palatoquadrate dorsally and the Meckelian cartilage ventrally. The gill arches (green) are situated internally to the gill filaments, and the nasal capsules (blue) open to the exterior by means of paired nostrils. (After Janvier 1996.)
There are numerous other characteristics of the soft anatomy and physiology (e.g. myelinated nerve fibres, sperms passing through urinary ducts, etc.), which are unique to the gnathostomes among extant craniates, but cannot by observed in fossils.
Discussion of Phylogenetic Relationships
Gnathostomes have long been placed as the sister-group of the ensemble of the Agnatha, or jawless vertebrates, regarded as a clade. Current phylogenies, however, suggest that the Agnatha are not a clade, and that, among Recent craniates, the gnathostomes are the sister-group of the Hyperoartia (lampreys) only. Among fossil craniates, the Osteostraci share the largest number of uniquely shared derived characteristics with the gnathostomes.
The controversies about gnathostome interrelationships mainly bear on the position of the two major fossil groups, the Placodermi and Acanthodii. Placoderms have long been regarded as the sister-group of chondrichthyans, with which they share an eye-stalk (a small cartilage linking the eyeball to the braincase) and possibly a pelvic clasper. It has also been suggested that they could be the sister-group of the osteichthyans, because they share with them large dermal plates and a median dermal bone in the palate, the parasphenoid. The current consensus is that placoderms are the sister-group to all other known gnathostomes. All other gnathostomes share a special arrangement of the eye muscles (the superior oblique eye muscle being attached anteriorly in the orbit), whereas that of placoderms is similar to that in the Osteostraci (the superior oblique eye muscle being attached posteriorly in the orbit). Also, the braincase of placoderms shows a separate ossification containing the nasal capsules, whereas the latter are always included in the braincase in all other gnathostomes.
The Acanthodii have long been regarded as closely related to either chondrichthyans or placoderms, but they are now almost unanimously regarded as the sister-group of the Osteichthyes. They share with them otoliths in the inner ear, lepidotrich-like scales on fins, and a narrow-based braincase with a transverse ventrale fissure. Acanthodians, however, are rarely known from articulated specimens, and their internal anatomy remains poorly known. It is possible that Acanthodii is paraphyletic.
References
Carroll, R. H. (1988). Vertebrate paleontology and evolution. W. H. Freeman & Co. New York.
Forey, P. L. (1980). Latimeria: a paradoxical fish. Proceedings of the Royal Society of London, B, 208, 369-384.
Gardiner, B. G. (1984). The relationships of placoderms. Journal of Vertebrate Paleontology, 4, 379-395.
Goujet, D. (1984). Placoderm interrelationships: a new interpretation, with a short review of placoderm classification. Proceedings of the Linnean Society of New South Wales. 107, 211-241.
Jarvik, E. (1977).The systematic position of acanthodian fishes. In Problems in vertebrate evolution (ed. S. M. Andrews, R. S. Miles and A. D. Walker), pp. 199-225. Academic Press, London.
Mallatt, J. (1984). Early vertebrate evolution: pharyngeal structure and the origin of gnathostomes. Journal of Zoology, 204, 169-183.
Miles, R. S. (1973). Relationships of acanthodians. In Interrelationships of fishes (ed. P. H. Greenwood, R. S. Miles, and C. Patterson). Zoological Journal of the Linnean Society, Supplement 1, 53, 63-103.
Schultze, H. P. (1993). Patterns of diversity in the skull of jawed fishes. In The skull (ed. J. Hanken and B. K. Hall), Vol. 2, pp. 189-254. University of Chicago Press.
Young, G. C. (1986). The relationships of placoderm fishes. Zoological Journal of the Linnean Society, 88, 1-57.
Information on the Internet
Deep homologies in the pharyngeal arches. Pharyngula.Title Illustrations

| Scientific Name | Hippopotamus amphibius |
|---|---|
| Location | Okavango Delta of Botswana |
| Sex | Male |
| Body Part | head |
| Copyright |
© 1997 Greg and Marybeth Dimijian

|
About This Page
Muséum National d'Histoire Naturelle Paris, France
Page copyright © 1997
 Page: Tree of Life
Gnathostomata. Jawed Vertebrates.
Authored by
Philippe Janvier.
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Gnathostomata. Jawed Vertebrates.
Authored by
Philippe Janvier.
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Citing this page:
Janvier, Philippe. 1997. Gnathostomata. Jawed Vertebrates. Version 01 January 1997 (under construction). http://tolweb.org/Gnathostomata/14843/1997.01.01 in The Tree of Life Web Project, http://tolweb.org/